НАДН гликолиза могут доставляться в митохондрии. Челночный механизм
НАДН гликолиза могут доставляться в митохондрии
Молекулы НАДН, образованные в шестой реакции гликолиза, в зависимости от наличия кислорода имеют, как минимум, два пути своего дальнейшего превращения:
- либо остаться в цитозоле и вступить в одиннадцатую реакцию гликолиза (анаэробные условия),
- либо проникнуть в митохондрию и окислиться в дыхательной цепи (аэробные условия).
Челночные системы
Так как сама молекула НАДН через мембрану не проходит, то существуют специальные системы, принимающие атомы водорода от НАДН в цитоплазме и отдающие их в матриксе митохондрий. Эти системы получили название челночные системы.
Определены две основные челночные системы – глицеролфосфатная и малат-аспартатная.
На основании наличия алкогольдегидрогеназы во многих тканях, в том числе и в нервной, дискутируется вопрос о существовании этанол-ацетальдегидной челночной системы, однако однозначных экспериментальных доказательств пока не получено.
Глицеролфосфатный челночный механизм
Ключевыми ферментами глицеролфосфатного челнока являются изоферменты глицерол-3-фосфат-дегидрогеназы – цитоплазматический и митохондриальный. Они отличаются своими коферментами: у цитоплазматической формы – НАД, у митохондриальной – ФАД.
В цитозоле метаболиты гликолиза – диоксиацетонфосфат и НАДН образуют глицерол-3-фосфат, поступающий в матрикс митохондрий. Там он окисляется с образованием ФАДН2. Далее ФАДН2 направляется в дыхательную цепь и используется для получения энергии. Таким образом, в результате действий челнока цитозольный НАДН+H+как бы "превращается" в митохондриальный ФАДН2.
Этот челнок активен в печени и белых скелетных мышцах и необходим для получения энергии из глюкозы при работе клетки.
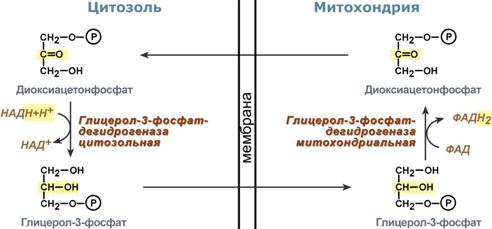 Схема работы глицерол-фосфатной челночной системы
Схема работы глицерол-фосфатной челночной системы Однако в адипоците и гепатоците в состоянии покоя и после еды часть глицерол-3 фосфата в митохондрию не пойдет, а будет использоваться в цитозоле для синтеза фосфолипидов и триацилглицеролов.
Малат-аспартатный челночный механизм
Ключевыми ферментами этого челнока являются изоферменты малатдегидрогеназы – цитоплазматический и митохондриальный. Он является распространенным по всем тканям.
Этот механизм более сложен: постоянно идущие в цитоплазме при участии фермента аспартатаминотрансферазы (АСТ) реакции трансаминирования аспарагиновой кислоты с α-кетоглутаратом поставляют оксалоацетат, который под действием цитозольного пула малатдегидрогеназы и за счет "гликолитического" НАДН восстанавливается до яблочной кислоты (малата).
Последняя антипортом с α-кетоглутаратом проникает в митохондрии и, являясь метаболитом ЦТК, окисляется в оксалоацетат с образованием НАДН. Так как мембрана митохондрий непроницаема для оксалоацетата, то он при помощи аспартатаминотрансферазы трансаминируется до аспарагиновой кислоты, которая в обмен на глутамат выходит в цитозоль.
Таким образом, атомы водорода от цитозольного НАДН перемещаются в состав митохондриального НАДН.
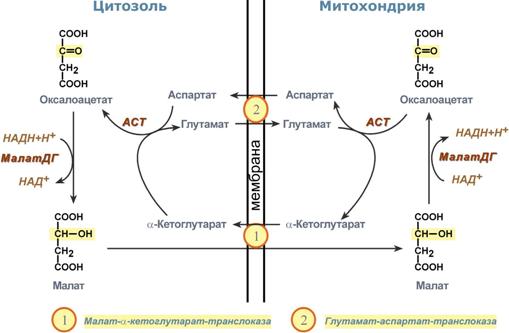 Схема работы малат-аспартатной челночной системы
Схема работы малат-аспартатной челночной системыВы можете спросить или оставить свое мнение.
biokhimija.ru
Челночные механизмы переноса водорода
Никотинамидные дегидрогеназы находятся не только в матриксе митохондрий, но и в цитозоле. Митохондриальная мембрана непроницаема для НАД, поэтому НАДН2, который образуется в ЦП, может передать свой водород в митохондрию только с помощью специальных субстратных ЧЕЛНОЧНЫХ МЕХАНИЗМОВ. В митохондрию из ЦП передается не сам НАДН2, а только водород, отнятый от него. Переносимый водород включается в молекулу вещества-челнока, способного проникать через митохондриальную мембрану. В митохондрии вещество-челнок отдает водород на митохондриальный НАД или ФАД и возвращается обратно в цитозоль.
В клетках организма человека существуют 2 типа челночных механизмов.
1. МАЛАТ-АСПАРТАТНЫЙ челнок (наиболее универсален для клеток организма). С высокой скоростью работает в миокарде, почечной ткани, печени.
В этой транспортной системе водород от цитоплазматического НАД передается на митохондриальный НАД(!), поэтому в митохондриях образуется 3 молекулы АТФ и не происходит потери энергии при переносе водорода. Для ткани печени малат-аспартатная система особенно важна, так как из митохондрии выводится Ацетил-КоА (в виде цитрата), а водород попадает в митохондрию (в составе малата).
Таким образом, происходит не только челночный транспорт водорода от цитоплазматического НАД к митохондриальному, а и обратный транспорт Ацетил-КоА из митохондрий в цитоплазму в виде цитрата. В цитоплазме Ацетил-КоА может быть использован для синтеза жирных кислот.
ЩУК может вернуться в цитоплазму и другим способом: она может вступить в реакцию трансаминирования с глутаминовой кислотой (СМОТРИТЕ РИСУНОК)
2. ГЛИЦЕРОФОСФАТНЫЙ челнок (встречается реже).
Вэтой транспортной системе водород от цитоплазматического НАД передается на митохондриальный ФАД(!), и в митохондриях образуется 2 молекулы АТФ вместо 3-х - происходит потеря энергии при переносе водорода.
В клетке существует не только челночный транспорт водорода от цитоплазматического НАД к митохондриальному. Происходит и обратный транспорт Ацетил-КоА из митохондрий в цитоплазму в виде цитрата. В цитоплазме Ацетил-КоА может быть использован для синтеза жирных кислот.
studfiles.net
Челночные системы
Строение и обмен углеводов | 26 |
|
|
Иллюстрацией к сказанному служит отличие |
|
изоферментов лактатдегидрогеназ (ЛДГ) друг |
|
от друга. Сердечный изофермент ЛДГ-1обла- |
|
дает высоким сродством к молочной кислоте и |
|
"стремится" поднять концентрацию пирувата с |
|
целью его включения в ЦТК и получения |
|
энергии для деятельности миокарда. Большое |
|
количество митохондрий и поступление сюда |
|
лактата из других органов обеспечивает рабо- |
|
ту сердца при аэробных условиях. При не- |
|
хватке кислорода свойства ЛДГ-1не изменят- | • |
ся, он по-прежнемубудет сдвигать реакцию в |
|
сторону продукции пировиноградной кислоты, и накопление пирувата вызовет необратимые повреждения кардиомиоцита.
В тоже время изофермент скелетной мышцы ЛДГ-5обладает высоким сродством к пирувату, при отсутствии кислорода в клетке быстро и эффективно превращает его в лактат, легко проникающий сквозь мембраны и удаляемый из мышцы.
Таким образом, в анаэробных условиях сильнее будет страдать сердечная мышца, что, собственно говоря, и наблюдается в медицинской практике.
Важно то, что пировиноградная кислота является для клетки токсичным веществом, и клетке необходимо избавиться от нее каким угодно образом. Так как через мембраны она не проходит, то обезвреживание достигается превращением пирувата 1) в лактат; 2) в ацетил-S-КоА;3) в аланин (см "Аланинаминотрансфераза"), 4) в оксалоацетат.
Молекулы НАДН, образованные в шестой реакции гликолиза, в зависимости от наличия кислорода имеют, как минимум, два пути своего дальнейшего превращения:
oлибо остаться в цитозоле и вступить в лактатдегидрогеназную реакцию (анаэробные условия),
o либо проникнуть в митохондрию и окислиться в дыхательной цепи (аэробные условия), Так как сама молекула НАДН через митохондриальную мембрану не проходит, то су-
ществуют системы, принимающие этот водород в цитоплазме и отдающие его в матриксе митохондрий. Они называются челночные системы.
Определены две основные челночные системы – глицеролфосфатная ималат-
аспартатная.
Глицеролфосфатный челночный механизм
Ключевыми ферментами глицеролфосфатного челнока являются изоферменты глице- рол-3-фосфат-дегидрогеназы – цитоплазматический и митохондриальный. Они отличаются своими коферментами: у цитоплазматической формы – НАД, у митохондриальной – ФАД.
В цитозоле метаболиты гликолиза – диоксиацетонфосфат и НАДН образуют глицерол- 3-фосфат,поступающий в матрикс митохондрий. Там он окисляется с образованием ФАДН2. Далее ФАДН2 направляется в дыхательную цепь и используется для получения энергии.
Работа глицеролфосфатного челночного механизма актуальна при необходимости получить энергию из глюкозы при работе клетки.
Однако, если в клетке имеется избыток энергии (состояние покоя,после еды) то гли-церол-3фосфат будет использоваться в цитозоле для синтеза жиров (см "Синтез триацилглицеролов").
biokhimija.ru | Тимин О.А. Лекции по биологической химии | 27 |
|
|
|
Малат-аспартатныйчелночный механизм
Этот механизм более сложен: постоянно идущие в цитоплазме реакции трансаминирования аспартата поставляют оксалоацетат, который под действием цитозольного пула малатдегидрогеназы и "гликолитического" НАДН восстанавливается до яблочной кислоты. Последняя антипортом с α-кетоглутаратомпроникает в митохондрии и, являясь метаболитом ЦТК, окисляется в оксалоацетат с образованием НАДН. Так как мембрана митохондрий непроницаема для оксалоацетата, то он аминируется до аспарагиновой кислоты, которая в обмен на глутамат выходит в цитозоль.
studfiles.net
- челночные системы - Биохимия
Молекулы НАДН, образованные в шестой реакции гликолиза, в зависимости от наличия кислорода имеют, как минимум, два пути своего дальнейшего превращения:
- либо остаться в цитозоле и вступить в одиннадцатую реакцию гликолиза (анаэробные условия),
- либо проникнуть в митохондрию и окислиться в дыхательной цепи (аэробные условия).
Челночные системы
Так как сама молекула НАДН через мембрану не проходит, то существуют специальные системы, принимающие атомы водорода от НАДН в цитоплазме и отдающие их в матриксе митохондрий. Эти системы получили название челночные системы.
Определены две основные челночные системы – глицеролфосфатная и малат-аспартатная.
На основании наличия алкогольдегидрогеназы во многих тканях, в том числе и в нервной, дискутируется вопрос о существовании этанол-ацетальдегидной челночной системы, однако однозначных экспериментальных доказательств пока не получено.
Глицеролфосфатный челночный механизм
Ключевыми ферментами глицеролфосфатного челнока являются изоферменты глицерол-3-фосфат-дегидрогеназы – цитоплазматический и митохондриальный. Они отличаются своими коферментами: у цитоплазматической формы – НАД, у митохондриальной – ФАД.
В цитозоле метаболиты гликолиза – диоксиацетонфосфат и НАДН образуют глицерол-3-фосфат, поступающий в матрикс митохондрий. Там он окисляется с образованием ФАДН2. Далее ФАДН2 направляется в дыхательную цепь и используется для получения энергии. Таким образом, в результате действий челнока цитозольный НАДН+H+как бы "превращается" в митохондриальный ФАДН2.
Этот челнок активен в печени и белых скелетных мышцах и необходим для получения энергии из глюкозы при работе клетки.
Схема работы глицерол-фосфатной челночной системы Однако в адипоците и гепатоците в состоянии покоя и после еды часть глицерол-3 фосфата в митохондрию не пойдет, а будет использоваться в цитозоле для синтеза фосфолипидов и триацилглицеролов.
Малат-аспартатный челночный механизм
Ключевыми ферментами этого челнока являются изоферменты малатдегидрогеназы – цитоплазматический и митохондриальный. Он является распространенным по всем тканям.
Этот механизм более сложен: постоянно идущие в цитоплазме при участии фермента аспартатаминотрансферазы (АСТ) реакции трансаминирования аспарагиновой кислоты с α-кетоглутаратом поставляют оксалоацетат, который под действием цитозольного пула малатдегидрогеназы и за счет "гликолитического" НАДН восстанавливается до яблочной кислоты (малата).
Последняя антипортом с α-кетоглутаратом проникает в митохондрии и, являясь метаболитом ЦТК, окисляется в оксалоацетат с образованием НАДН. Так как мембрана митохондрий непроницаема для оксалоацетата, то он при помощи аспартатаминотрансферазы трансаминируется до аспарагиновой кислоты, которая в обмен на глутамат выходит в цитозоль.
Таким образом, атомы водорода от цитозольного НАДН перемещаются в состав митохондриального НАДН.
Схема работы малат-аспартатной челночной системыВы можете спросить или оставить свое мнение.
biokhimija.ru
Челночные системы
Челночные системы
Так как сама молекула НАДН через мембрану не проходит, то существуют специальные системы, принимающие атомы водорода от НАДН в цитоплазме и отдающие их в матриксе митохондрий. Эти системы получили название челночные системы.
Определены две основные челночные системы – глицеролфосфатная и малат-аспартатная.
На основании наличия алкогольдегидрогеназы во многих тканях, в том числе и в нервной, дискутируется вопрос о существовании этанол-ацетальдегидной челночной системы, однако однозначных экспериментальных доказательств пока не получено.
.
Глицеролфосфатный челночный механизм
Ключевыми ферментами глицеролфосфатного челнока являются изоферменты глицерол-3-фосфат-дегидрогеназы – цитоплазматический и митохондриальный. Они отличаются своими коферментами: у цитоплазматической формы – НАД, у митохондриальной – ФАД.
В цитозоле метаболиты гликолиза – диоксиацетонфосфат и НАДН образуют глицерол-3-фосфат, поступающий в матрикс митохондрий. Там он окисляется с образованием ФАДН2. Далее ФАДН2 направляется в дыхательную цепь и используется для получения энергии. Таким образом, в результате действий челнока цитозольный НАДН+H+ как бы "превращается" в митохондриальный ФАДН2.
Этот челнок активен в печени и белых скелетных мышцах и необходим для получения энергии из глюкозы при работе клетки.
Схема работы глицерол-фосфатной челночной системы
Однако, если в клетке имеется избыток энергии (состояние покоя, после еды), то часть глицерол-3 фосфата в митохондрию не пойдет, а будет использоваться в цитозоле гепатоцитов для синтеза фосфолипидов и триацилглицеролов.
.
Малат-аспартатный челночный механизм
Ключевыми ферментами этого челнока являются изоферменты малатдегидрогеназы – цитоплазматический и митохондриальный. Он является распространенным по всем тканям.
Этот механизм более сложен: постоянно идущие в цитоплазме реакции трансаминирования аспарагиновой кислоты поставляют оксалоацетат, который под действием цитозольного пула малатдегидрогеназы и за счет "гликолитического" НАДН восстанавливается до яблочной кислоты (малата).
Последняя антипортом с α-кетоглутаратом проникает в митохондрии и, являясь метаболитом ЦТК, окисляется в оксалоацетат с образованием НАДН. Так как мембрана митохондрий непроницаема для оксалоацетата, то он аминируется до аспарагиновой кислоты, которая в обмен на глутамат выходит в цитозоль.
Таким образом, атомы водорода от цитозольного НАДН перемещаются в состав митохондриального НАДН.
studfiles.net
Челночные системы
biokhimija.ru | Тимин О.А. Лекции по биологической химии | 203 |
|
|
|
Иллюстрацией к сказанному служит отличие изоферментов лактатдегидрогеназ (ЛДГ) друг от друга. Сердечный изофермент ЛДГ-1обладает высоким сродством к молочной кислоте и "стремится" поднять концентрацию пирувата с целью его включения в ЦТК и получения энергии для деятельности миокарда. Большое количество митохондрий и поступление сюда лактата из других органов обеспечивает работу сердца при аэробных условиях. При нехватке кислорода свойстваЛДГ-1не изменятся, онпо-прежнемубудет сдвигать реакцию в
сторону продукции пировиноградной кислоты, и накопление пирувата вызовет необратимые повреждения кардиомиоцита.
В тоже время изофермент скелетной мышцы ЛДГ-5обладает высоким сродством к пирувату, при отсутствии кислорода в клетке быстро и эффективно превращает его в лактат, легко проникающий сквозь мембраны и удаляемый из мышцы.
Таким образом, в анаэробных условиях сильнее будет страдать сердечная мышца, что, собственно говоря, и наблюдается в медицинской практике.
Важно то, что пировиноградная кислота является для клетки токсичным веществом, и клетке необходимо избавиться от нее каким угодно образом. Так как через мембраны она не проходит, то обезвреживание достигается превращением пирувата 1) в лактат; 2) в ацетил-S-КоА;3) в аланин (см "Аланинаминотрансфераза"), 4) в оксалоацетат.
Молекулы НАДН, образованные в шестой реакции гликолиза, в зависимости от наличия кислорода имеют, как минимум, два пути своего дальнейшего превращения:
oлибо остаться в цитозоле и вступить в лактатдегидрогеназную реакцию (анаэробные условия),
o либо проникнуть в митохондрию и окислиться в дыхательной цепи (аэробные условия), Так как сама молекула НАДН через митохондриальную мембрану не проходит, то су-
ществуют системы, принимающие этот водород в цитоплазме и отдающие его в матриксе митохондрий. Они называются челночные системы.
Определены две основные челночные системы – глицеролфосфатная ималат-
аспартатная.
Глицеролфосфатный челночный механизм
Ключевыми ферментами глицеролфосфатного челнока являются изоферменты глице- рол-3-фосфат-дегидрогеназы – цитоплазматический и митохондриальный. Они отличаются своими коферментами: у цитоплазматической формы – НАД, у митохондриальной – ФАД.
В цитозоле метаболиты гликолиза – диоксиацетонфосфат и НАДН образуют глицерол- 3-фосфат,поступающий в матрикс митохондрий. Там он окисляется с образованием ФАДН2. Далее ФАДН2 направляется в дыхательную цепь и используется для получения энергии.
Работа глицеролфосфатного челночного механизма актуальна при необходимости получить энергию из глюкозы при работе клетки.
Однако, если в клетке имеется избыток энергии (состояние покоя,после еды) то гли-церол-3фосфат будет использоваться в цитозоле для синтеза жиров (см "Синтез триацилглицеролов").
Строение и обмен углеводов | 204 |
|
|
Малат-аспартатныйчелночный механизм
Этот механизм более сложен: постоянно идущие в цитоплазме реакции трансаминирования аспартата поставляют оксалоацетат, который под действием цитозольного пула малатдегидрогеназы и "гликолитического" НАДН восстанавливается до яблочной кислоты. Последняя антипортом с α-кетоглутаратомпроникает в митохондрии и, являясь метаболитом ЦТК, окисляется в оксалоацетат с образованием НАДН. Так как мембрана митохондрий непроницаема для оксалоацетата, то он аминируется до аспарагиновой кислоты, которая в обмен на глутамат выходит в цитозоль.
studfiles.net
Челночные механизмы переноса водорода
Никотинамидные дегидрогеназы находятся не только в матриксе митохондрий, но и в цитозоле. Митохондриальная мембрана непроницаема для НАД, поэтому НАДН2, который образуется в цитозоле, может передать свой водород в митохондрию только с помощью специальных субстратных ЧЕЛНОЧНЫХ МЕХАНИЗМОВ. В митохондрию из цитозоля передается не сам НАДН2, а только водород, отнятый от него. Переносимый водород включается в молекулу вещества-челнока, способного проникать через митохондриальную мембрану. В митохондрии вещество-челнок отдает водород на митохондриальный НАД или ФАД и возвращается обратно в цитозоль.
В клетках организма человека существуют 2 типа челночных механизмов.
1. МАЛАТ-АСПАРТАТНЫЙ челнок (наиболее универсален для клеток организма). С высокой скоростью работает в миокарде, почечной ткани, печени.
В этой транспортной системе водород от цитоплазматического НАД передается на митохондриальный НАД(!), поэтому в митохондриях образуется 3 молекулы АТФ и не происходит потери энергии при переносе водорода. Для ткани печени малат-аспартатная система особенно важна, так как из митохондрии выводится Ацетил-КоА (в виде цитрата), а водород попадает в митохондрию (в составе малата).
Таким образом, происходит не только челночный транспорт водорода от цитоплазматического НАД к митохондриальному, а и обратный транспорт Ацетил-КоА из митохондрий в цитоплазму в виде цитрата. В цитоплазме Ацетил-КоА может быть использован для синтеза жирных кислот.
ЩУК может вернуться в цитоплазму и другим способом: она может вступить в реакцию трансаминирования с глутаминовой кислотой (СМОТРИТЕ РИСУНОК)
2. ГЛИЦЕРОФОСФАТНЫЙ челнок (встречается реже).
Вэтой транспортной системе водород от цитоплазматического НАД передается на митохондриальный ФАД(!), и в митохондриях образуется 2 молекулы АТФ вместо 3-х - происходит потеря энергии при переносе водорода.
В клетке существует не только челночный транспорт водорода от цитоплазматического НАД к митохондриальному. Происходит и обратный транспорт Ацетил-КоА из митохондрий в цитоплазму в виде цитрата. В цитоплазме Ацетил-КоА может быть использован для синтеза жирных кислот.
studfiles.net







